Enterococcus faecium NRRL B-2354: Tempest in a Teapot or Serious Foodborne Pathogen?
A particular bacterial strain, now known as Enterococcus faecium NRRL B-2354, has been used within the food industry for a variety of useful purposes for perhaps the past 64 years. It is known under various names such as ATCC 8459, NRRL B-2354 and NCIMB 2699. It was also previously labeled IFO3778, NCDO989, NCIB 2699 and NCTC 2699.[1,2] However, these last four strain designations no longer appear to be available. Pathogenic microorganisms have been grouped into four categories based upon risk: biosafety level (BSL) 1, 2, 3 and 4. Recently, the American Type Culture Collection (ATCC) listed all E. faecium strains as BSL2 pathogens. This is the same category into which Salmonella and Listeria monocytogenes are grouped. This has caused concern within various sectors of the food industry, which have used this particular strain (e.g., NRRL B-2354) for many decades. BSL1 organisms are regarded as nonpathogenic. Other culture collections referencing the same strain have not done so.
NRRL B-2354 should continue to be regarded as “a heat resistant, spoilage, nonpathogenic organism that has been used as a test organism for studying the destruction of bacteria…in a food pilot plant.”[3]
Background and Need for E. faecium NRRL B-2354
Prevention is the focus of the Food Safety Modernization Act.[4] In particular, section 103 indicates that Chapter IV (21 U.S.C. 341 et seq.) is amended by adding section 418 “Hazard Analysis and Risk-based Preventive Controls,” which includes subsection “(c) Preventive Controls.” This in part states that “the owner, operator, or agent in charge of a facility shall identify and implement preventive controls, including at critical control points...” All food manufacturers will need to conduct a hazard analysis, the purpose of which is to identify and implement documented preventative control measures. Consistent with that approach is the validation of thermal processes, many of which are too dynamic to adequately model in a laboratory setting. Thermal processing of foods can involve drying products, thus lowering their water activity (a{w}) as their temperatures increase throughout the process. It is well documented that the heat resistance of microbial populations increases as a{w} decreases.[5] In addition, microbial lethality should increase as temperature increases, all things being equal. However, all things are not equal. These two principles act counter to one another with regard to microbiological heat resistance, resulting in the inability to accurately predict the microbicidal efficacy of many thermal processes. This is illustrated at right with Salmonella and with E. faecium NRRL B-2354, wherein more lethality occurred in a wheat-based product at lower temperatures and higher a{w} than at much higher temperatures and lower a{w}. This information was presented at the 2006 International Association for Food Protection annual meeting and is referenced in a forthcoming 2012 publication (see Figures 1 and 2).[6,7]
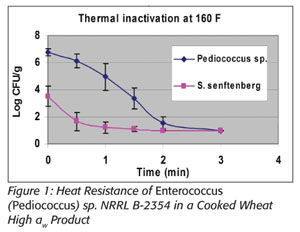
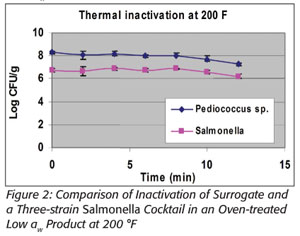
Furthermore, it is not correct to extrapolate thermal heat resistance between different food matrices even when they are at the same water activities.[5] Different approaches to modeling microbial heat resistance exist, but these present many potential limitations. Hence, there is a need for an appropriate surrogate microbe to validate actual in-factory conditions. Some researchers believe that E. faecium NRRL B-2354 is the only appropriate surrogate for use in thermal processing of dry foods in place of Salmonella.[8]
I have[9,10] described surrogates as harmless microbes with correlated survival and growth parameters to specific pathogens that are frequently used to validate the efficacy of critical limits for Critical Control Points. They can be naturally occurring or artificially added and should be nonpathogenic (emphasis mine). Both the U.S. Department of Agriculture (USDA) and Food and Drug Administration have acknowledged the use of surrogate microorganisms as a valid approach where appropriate. An advantage of using surrogates is that they can be used under actual factory or pilot plant conditions to validate the efficacy of a process without the need to attempt to simulate such processes in a laboratory.[10]
Identity of NRRL B-2354
Data in a study by Ma et al.[11] indicated that rDNA sequencing (500 base-pair region) performed on ATCC 8459 indicated it to be in the genus Enterococcus. However, further speciation by this technique was not possible. It appeared closely related to E. faecium, or E. hirae or E. durans.[11] The authors referred to it in their paper as an E. faecium based upon further phenotypic (e.g., Vitek) analysis. However, Franz et al.[12] stated that “The classical taxonomy of the enterococci is vague because there are no phenotypic characteristics that unequivocally distinguish them from other Gram-positive, catalase negative, coccus-shaped bacteria.” Hence, the identification of this strain by that means should not be considered conclusive, especially as the differences between the likely species referenced above via rDNA analysis were on the order of one to two base pairs.[13] The determining factor was a biochemical analysis with a Vitek unit, which is a biochemically based phenotypic approach that is dependent upon the database of patterns stored within each unit. Results are not reproducible in a laboratory apart from use of another Vitek unit with the same database. Hence, the identification described by Ma et al. could be viewed as indeterminate. The consequences of deciding species identity of this organism are more than a purely academic exercise, since the species durans and hirae have rarely, if ever, been associated with human illness. Nevertheless, the work of Ma et al. appears to have been used to call the strain E. faecium by one or more of the culture collections and is referenced as a definitive confirmation by the Almond Board of California.[14]
It is noteworthy that ATCC does not guarantee the accuracy of the strain identity. A note at the bottom of the link on the ATTC website wherein ATCC 8459 is described reads, “While ATCC uses reasonable efforts to include accurate and up-to-date information on this site, ATCC makes no warranties or representations as to its accuracy. Citations from scientific literature and patents are provided for informational purposes only. ATCC does not warrant that such information has been confirmed to be accurate.”[1] The Almond Board of California specifically recommended the use of the NRRL B-2354 strain as it was provided without cost and had been used in a surrogate study.[14]
History of Use in the Food Industry
Other Enterococcus strains: The good, the bad and the ugly
Historically, tests for Enterococcus spp. in foods or food processing environments or surfaces have been used as indicators of hygiene, fecal contamination, quality indicators or sanitation efficacy. A review of the beneficial uses of Enterococcus spp. (e.g., bacteriocins, probiotics, cheese manufacture) and their implication in food spoilage and as fecal hygienic indicators and in disease was provided by Franz et al.[12] The authors demonstrated that contamination of cheeses and ready-to-eat meats is fairly common. However, foodborne infections with Enterococci are unproven. Food microbiologists typically regard Enterococcus spp. to be food quality indicators, or spoilage organisms (in some foods), but not food pathogens. This is despite speculation and research about their role in foodborne illness over many decades. Enterococci were not listed as food pathogens in a number of texts,[15–17] nor were they found in any of the International Commission on Microbiological Specifications for Foods lists of moderate, serious or severe foodborne pathogens. Franz et al. states, “E. faecalis and E. faecium have been suspected, but remain unconfirmed as causative agents of foodborne illness.” The same is still true today. Correspondence with an Enterococcus spp. expert at the Centers for Disease Control and Prevention (CDC)[18] indicated that she “…did not think there’s any new knowledge or newly recognized risk to food microbiologists.” The author acknowledged that in other contexts, E. faecium is a recognized human pathogen and went on to indicate that ATCC had likely characterized this strain (and all other E. faecium strains in their collection) as BSL2 pathogens out of an abundance of caution.
In my view, it makes sense for some strains of E. faecium to be classified as BSL2 organisms, but not all strains, as will be further demonstrated below. Virulence factors with some strains of E. faecalis have been identified, but those for E. faecium are less understood.[12]
Nevertheless, Enterococci are common causes of hospital-acquired infections (as are generic Escherichia coli, some of which have been recommended as surrogates in other contexts).[19] In my view, those described in that reference would not be appropriate for thermal process validation as surrogates for Salmonella. Some researchers have shown apparent strain identity in selected strains of Enterococci in meat products and in humans, from which they inferred a foodborne route of entry.[20,21] Wegener[20] also isolated vancomycin-resistant Enterococci from foods. Others have inferred a food route from some animal feeding studies in which diarrhea occurred after feeding.[22] The literature clearly shows that Enterococci can cause illness as an opportunistic infectious agent, although foodborne infection or intoxication has never been proven.
It is likely that there are also species and strain-to-strain differences in pathogenicity within the Enterococcus species faecium. It is significant that a strain of E. faecium was used as a probiotic with humans and found to reduce cholesterol in at least two studies,[23,24] and one preparation (E. faecium cernell 68, also known as E. faecium SF68) was shown to suppress E. coli-mediated diarrhea in gnotobiotic pigs.[12,25] I am also aware of studies in the 1940s in which E. faecalis, a related organism, was used as a starter culture in cheese.[26–29] Cheese and other foods containing high populations of this E. faecalis as well as broth cultures were ingested by human volunteers with no illness recorded.[30] Several other alleged “food poisoning” strains of E. faecalis were also ingested by humans in high numbers and various types of foods with little effect. In another study, E. faecium strains were isolated from human breast milk.[31] The strains were screened for virulence factors. The authors concluded, “These findings suggest that milk of healthy mothers may be a source of avirulent E. faecium isolates to the newborns.”
E. faecium NRRL B-2354 (ATCC 8459, NCIMB 2699) Uses in the Food Industry
Originally, this organism was called Micrococcus freudenreichii ATCC 8459. The taxonomic name M. freudenreichii was eventually dropped[32]. Previously, it was “species incertae sedis.”[33] It is my understanding that the strain was dropped because the G+C content differed too much from other members of the genus Micrococcus. The strain was later called (unofficially) Pediococcus sp. NRRL B2354 by USDA3 until the early part of the past decade, when the ATCC began to refer to it as E. faecium ATCC 8459. Sometime late in the last decade, ATCC elected to call all strains of E. faecium BSL2 organisms. This is unfortunate for the food industry for a variety of reasons: ATCC 8459 has been used as an adjunct starter culture for cheese manufacture[34] and also as a surrogate in place of food pathogens (like Salmonella and L. monocytogenes) to validate the safety of thermal processing to kill pathogens in a variety of foods. In addition, this same strain, also known as NCIB 2699, was noted to produce a bacteriocin active against L. monocytogenes,[35,36] thus showing potential for further use of the organism in the manufacture of safe foods.
I have seen no evidence in the literature to suggest this particular strain represents a health risk in food, but much evidence of beneficial use in food development and research for the following reasons.
1. The microbe formerly known as M. freudenreichii (not necessarily ATCC 8459) was commonly found in foods in the past. Consequently, if this microbe was a pathogen, it is likely that we would have discovered it previously. The use of M. freudenreichii (not necessarily ATCC strain 8459) as an adjunct starter culture was also previously documented.[37–42]
2. The microbe (not necessarily ATCC 8459) has been shown to contribute significantly to the ripening process of various cheeses.[38,43] Fox and Tobin[43] stated that “adjunct bacteria, including mesophilic and thermophilic lactobacilli, M. freudenreichii and Pediococcus pentosaceus have shown some promise in the manufacture of low-fat cheese with reduced defects and improved flavor.” Please note: Adjunct bacteria are deliberately added to cheese.
3. Three patents have been issued for its deliberate use in cheese manufacture for the purpose of enhancing the flavor as adjunct cultures.[34, 44–47] Luksas44 states that the cheese is “Inoculated with a nontoxic Micrococcus such as…Micrococcus freudenreichii ATCC 407 or 8459.” This suggests an industrial food use of the specific strain for the last 40 years. Bhomick and Marth46 screened cultures for their proteolytic activity to find Micrococcus cultures that may be used to enhance cheese flavor development, including Micrococcus sp. ATCC 8459 and cited the work of Alford and Frazer,[47] who used M. freudenreichii to accelerate the ripening of Cheddar cheese.
4. M. freudenreichii has been used for at least 64 years by microbiologists as a nonpathogenic surrogate organism in place of pathogens to validate thermal processes of foods beginning, apparently with Speck[48] with regard to pasteurized milk and ice cream mix, Speck and Lucas[49] with chocolate milk and Collins et al.[50] with pasteurized milk. Other researchers have used this organism to study the heat resistance in concentrated ultrafiltered milk retentates,[51] juice products,[3] microwave processing of a variety of fluids,[52] apple cider,[53] oven treatment of wheat-based products,[6,7] cooking of ground beef[11] and almond roasting.[54]
Even if it is shown conclusively someday that some Enterococcus strains cause foodborne illness, in my view, we have a substantial track record of many decades of the use of ATCC 8459 (NRRL B-2354 or NCIMB 2699), and the M. freudenreichii MS66 strains[48] that provide no evidence of foodborne illness and a substantial public health benefit to its use as thermal surrogates in foods.
In addition, the Almond Board of California[14] recommended the use of this strain (NRRL B-2354, ATCC 8459) to validate the roasting of almonds. Since that time, other researchers have implicitly affirmed the safety of the organism, such as the paper by Jeong et al.,[54] the title of which is “Quantifying the performance of Pediococcus sp. (NRRL B-2354: Enterococcus faecium) as a nonpathogenic surrogate for Salmonella Enteritidis PT30 during moist-air convection heating of almonds” (emphasis mine). I am also aware of a number of companies in the United States who have done work or are now doing work in their pilot laboratories or actual production facilities with this organism.
In this author’s view, the BSL2 categorization of this organism by one culture collection (e.g., ATCC) will preclude the necessary thermal validation work that has gone on for 64 years and has contributed to the U.S. having the safest food supply in the world.
For the aforementioned reasons, I see no reason to back away from the statement in Annous et al.[3] (with reference to strain NRRL B-2354): This bacterium, originally isolated from milk and dairy utensils, “is a heat-resistant, spoilage, nonpathogenic organism that has been used as a test organism for studying the destruction of bacteria…in a food pilot plant.”
Implication of Enterococcus spp. in Human Foodborne Illness: Virulence Factors
Illnesses from Enterococcus species are significant sources of community- and hospital-acquired infections—in debilitated individuals, for example, those with underlying disease or who have received medical instrumentation, as indicated earlier. In this context, they have caused urinary tract infections, bacteremia, endocarditis, neonatal central nervous system infections and intra-abdominal and pelvic infections.[12] E. faecalis is isolated from the majority (80 percent) of human enterococcal infections, with the remainder mainly associated with E. faecium. The main concern about Enterococcus spp. (faecalis and faecium) in food has been with the potential for horizontal gene transfer of factors associated with virulence and antibiotic resistance.[12] A major focus of research related to Enterococcus spp. has been with those strains that have antibiotic resistance (particularly, but not limited to vancomycin resistance). This was highlighted with a volunteer feeding study in which an animal-derived Enterococcus strain with vancomycin resistance was transferred to a human strain within the intestinal tract of three of six human volunteers.[55] Hence, there is concern that introduction of an antibiotic-resistant strain could transfer such resistance to normal human flora (e.g., other Enterococcus spp. or related strains) that would then persist and result in a difficult-to-control infection should the patient become debilitated under some circumstances. However, although this potential exists, the human strain transformants were transient in the experiment.
Antibiotic Resistance and Virulence Factors of NRRL B-2354, ATCC 8459 and NCIMB 2699
The strain known as NCIMB 2699 that was deposited as ATCC 8459 with an accession date of January 8, 1978, appears to have no unusual antibiotic resistance. The site at www.ncimb.com/results.php indicates sensitivity to a number of antibiotics, including penicillin, streptomycin, chloramphenicol, tetracycline, polymyxin b and novobiocin. The site does not reference any antibiotic resistance associated with this strain. The site indicates that the strain was originally isolated from cheese.
Need for Further Research
This paper assumes that the strains represented to be the same in various culture collections have remained so, and mutations have not occurred. This assumption should be verified, better methods to confirm the identity of these strains should be employed and more comprehensive screening for antibiotic resistance should be performed.
Conclusion
It is my opinion that food manufacturers should not be discouraged from the use of E. faecium NRRL B-2354.
Jeffrey L. Kornacki, Ph.D., is president of Kornacki Microbiology Solutions Inc. in Madison, WI and is an adjunct assistant professor in the University of Georgia’s Department of Food Science.
13. Ma, L. 2007. Personal communication.
18. Anonymous. 2011. Correspondence with a CDC microbiologist.
Looking for quick answers on food safety topics?
Try Ask FSM, our new smart AI search tool.
Ask FSM →







