Meat Science Review
Sequence-specific removal of STECs via CRISPR-Cas9 system
Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) and CRISPR-associated proteins (Cas) together comprise the CRISPR-Cas system. In bacteria, this system confers an adaptive immunity against invading mobile elements such as viruses and plasmids. The CRISPR-Cas9 system has emerged as a programmable and versatile tool for precise genome editing in a wide variety of organisms. In contrast, research conducted in the Department of Animal Sciences at Colorado State University focused on exploiting the CRISPR-Cas9 system for selective killing of a bacterial pathogen relevant to meat safety by targeting specific virulence genes.
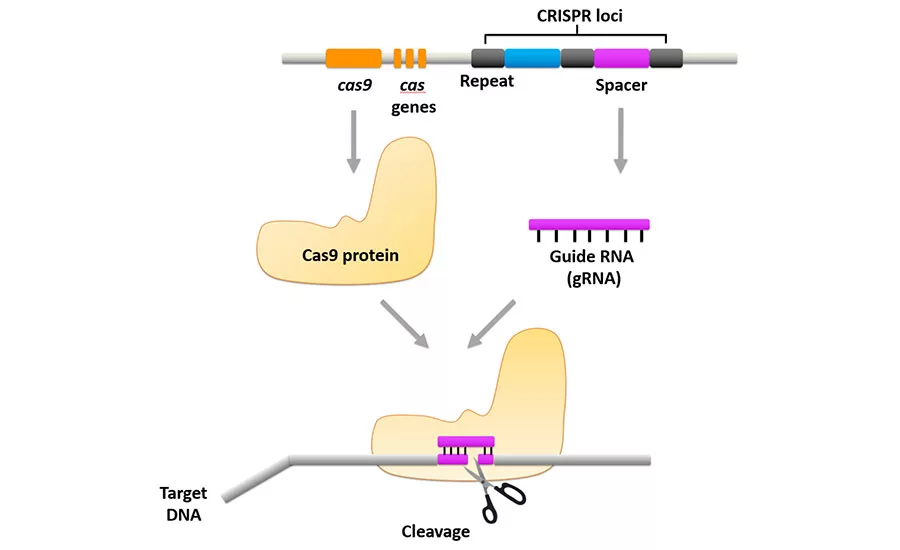
The CRISPR-Cas9 system contains two key factors: the Cas9 protein, which can cleave double-stranded DNA, and guide RNA (gRNA) which is transcribed from the CRISPR sequence (Figure 1). The function of the Cas9 protein is like a pair of scissors. Guide RNA is complimentary to the target sequence and is able to guide the Cas9 protein to the sequence-specific site of cleavage. The interaction between the Cas9 protein and the target DNA leads to the creation of a double-stranded DNA break. Such DNA cleavage can be used either to edit or to kill organisms. In recent studies, CRISPR-Cas9 systems have been explored for developing sequence-specific antimicrobials, which means that by targeting the cleavage of specific sites in a genome of interest, the kill should be limited to only organisms containing that specific gene or genes.
In this study, we designed and cloned gRNAs targeting Shiga toxin genes (stx1 or stx2) and used a two-plasmid platform to deliver this Shiga toxin-specific CRISPR-Cas9 system into bacterial cells to kill a Shiga toxin-producing Escherichia coli (STEC) strain. The CRISPR-Cas9 system can be programmed to selectively kill pathogens that harbor the Shiga toxin genes while leaving those non-target bacterial populations unaffected.
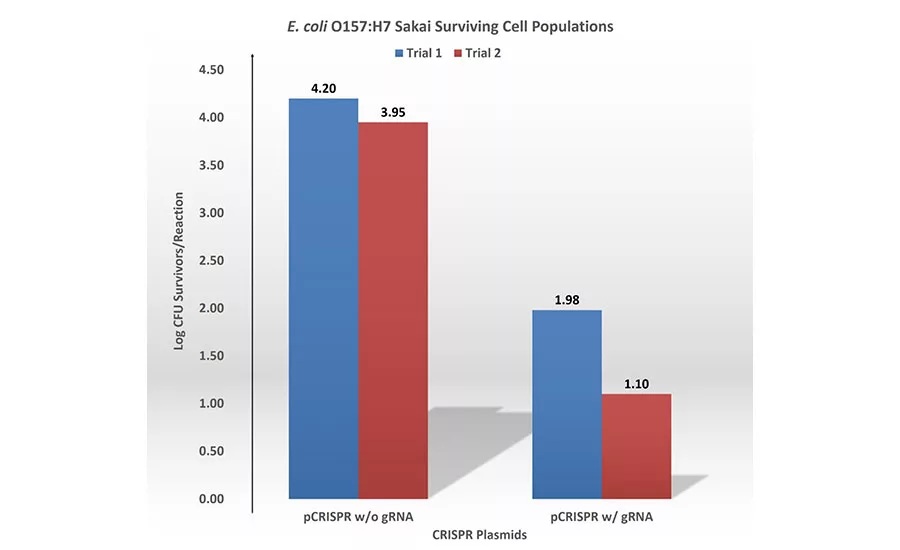
Shiga toxin gRNAs were designed by screening Shiga toxin gene sequences for NGG on the 3’ side. The designed 20-nucleotide gRNA was then cloned into a CRISPR plasmid (pCRISPR), resulting in a CRISPR plasmid with a gRNA (pCRISPR w/gRNA). The successful cloning of gRNA was confirmed by Sanger Sequencing. The Cas9 plasmid (pCas9) and pCRISPR with gRNA were then introduced into E. coli O157:H7 Sakai cells to determine the killing efficiency of the designed CRISPR-Cas9 system. In addition, pCas9 and pCRISPR without a gRNA were introduced into E. coli O157:H7 Sakai cells to serve as a control. When the pCRISPR with gRNA was introduced into the recipient E. coli O157:H7 cells containing pCas9 plasmids, an approximately 2.2- to 2.9-log lower number of E. coli O157:H7 cells was observed compared with that of the control pCRISPR without gRNA (Figure 2). This result provided evidence that introduction of a CRISPR-Cas9 system targeting Shiga toxin genes can achieve sequence-specific killing of STEC.
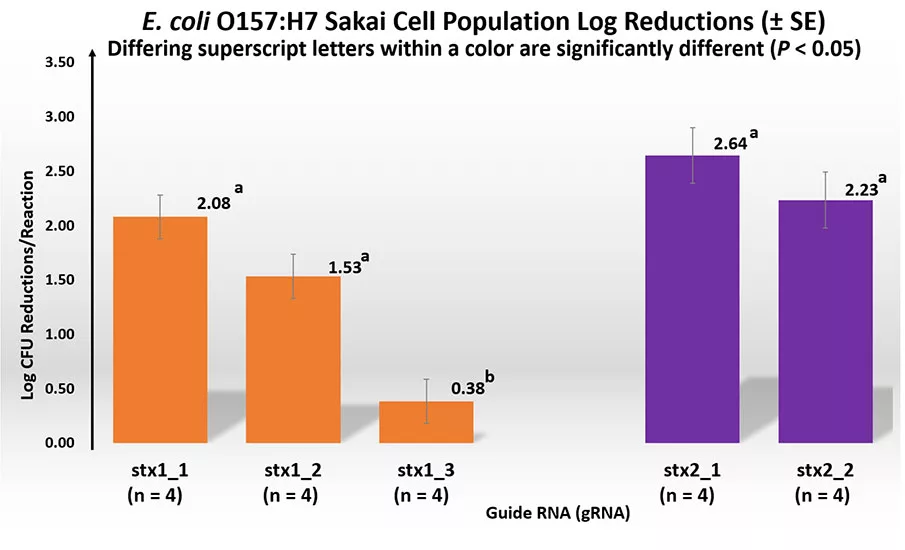
Killing target cells using the CRISPR-Cas9 system could be highly related to its gRNA sequence. In the next step, we designed three gRNAs (stx1-1, stx1-2 and stx1-3) that targeted the stx1 gene at different locations and two gRNAs (stx2-1 and stx2-2) that targeted the stx2 gene at different locations. We compared their efficiencies in killing E. coli O157:H7 Sakai cells. For the stx1 gene, we found no significant difference in cell reductions between cells that contained stx1-1 (2.08 log reduction) and stx1-2 (1.53 log reduction) gRNAs (Figure 3). However, significantly (P < 0.05) lower cell reductions were observed for cells containing the stx1-3 gRNA (0.38 log reduction) compared with the cells containing stx1-1 and stx1-2 gRNAs. For the stx2 gene, no significant (P > 0.05) differences in cell reductions were obtained between the stx2-1 (2.64 log reduction) and stx2-2 (2.23 log reduction) gRNAs (Figure 3). This result demonstrated varied reductions of E. coli O157:H7 Sakai cells when different gRNAs were used, suggesting that gRNAs are critical factors that determine the killing efficiencies of STEC.
In summary, this study provides proof-of-concept evidence that the CRISPR-Cas9 system with gRNA can selectively kill pathogenic bacteria by targeting a specific gene of choice. Furthermore, killing efficiencies can be improved by designing gRNAs to target virulence genes for sequence-specific removal of pathogenic bacteria, target antimicrobial resistance genes for killing antibiotic-resistant bacteria and target genes involved in biofilm development and formation of bacterial persister cells in biofilms. It also can improve sanitizer efficiency against biofilms in meat-processing environments. Further research will focus on developing a delivery system that allows us to apply the CRISPR-Cas9 system in real meat production and processing environments. NP
Citations
- Bielaszewska M, Schmidt H, Liesegang A, Prager R, Rabsch W, Tschäpe H, Cízek A, Janda J, Bláhová K, Karch H. 2000. Cattle can be a reservoir of sorbitol-fermenting Shiga toxin-producing Escherichia coli O157:H− strains and a source of human diseases. J Clin Microbiol, 38(9):3470-3.
- Bikard D, Euler CW, Jiang W, Nussenzweig PM, Goldberg GW, Duportet X, Fischetti VA, Marraffini LA. 2014. Exploiting CRISPR-Cas nucleases to produce sequence-specific antimicrobials. Nat Biotechnol, 32(11):1146-50. doi:10.1038/nbt.3043.
- Citorik RJ, Mimee M, Lu TK. 2014. Sequence-specific antimicrobials using efficiently delivered RNA-guided nucleases. Nat Biotechnol, 32(11):1141-5. doi:10.1038/nbt.3011.
- Doudna JA, Charpentier E. 2014. The new frontier of genome engineering with CRISPR-Cas9. Science, 346(6213), 1258096. doi:10.1126/science.1258096.
- Hsu PD, Lander ES, Zhang F. 2014. Development and applications of CRISPR-Cas9 for genome engineering. Cell, 157(6):1262-78. doi:10.1016/j.cell.2014.05.010.
- Jiang W, Bikard D, Cox D, Zhang F, Marraffini LA. 2013. RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nat Biotechnol, 31(3):233-9. doi: 10.1038/nbt.2508.
- Sternberg SH, Redding S, Jinek M, Greene EC, Doudna JA. 2014. DNA interrogation by the CRISPR RNA-guided endonuclease Cas9. Nature, 507(7490):62-7. doi:10.1038/nature13011.
- Wiedenheft B, Sternberg SH, Doudna JA. 2012. RNA-guided genetic silencing systems in bacteria and archaea. Nature, 482(7385):331-8. doi:10.1038/nature10886.
Looking for quick answers on food safety topics?
Try Ask FSM, our new smart AI search tool.
Ask FSM →







